Structural mechanism of the ATP-induced dissociation of rigor
myosin from actin.
Sebastian Kühner & Stefan Fischer*
Proceedings of the National
Academy
of Sciences USA (vol. 108, p. 7793-7798,
2011).
Abstract
Myosin is a true nanomachine, which
produces mechanical force from ATP hydrolysis by cyclically interacting
with actin filaments in a four-step cycle. The principle underlying
each step is that structural changes in separate regions of the protein
must be mechanically coupled. This sort of well defined coupling between parts of the motor
really is the essence of any type of motor (for instance the motions of
the valve and the piston are coupled in a car engine). The key to
understanding how myosin works as a nanomachine is to understand in
each step how the structural information is communicated from one part
of myosin to another (much like finding the crankshaft and the timing
belt in the engine analogy).
The step in which myosin dissociates from tightly bound actin (the
rigor state, thus called for
the stiffness of corpses when ATP gets depleted in muscle cells) is
triggered by the 30 Å distant binding of ATP.
Large conformational differences between the crystal structures make it
difficult to perceive the coupling mechanism of rigor dissociation. Here,
energetically accessible transition
pathways computed at atomic detail (using the
Conjugate Peak Refinement (CPR) method)
reveal a simple coupling mechanism for the reciprocal binding of ATP
and actin.
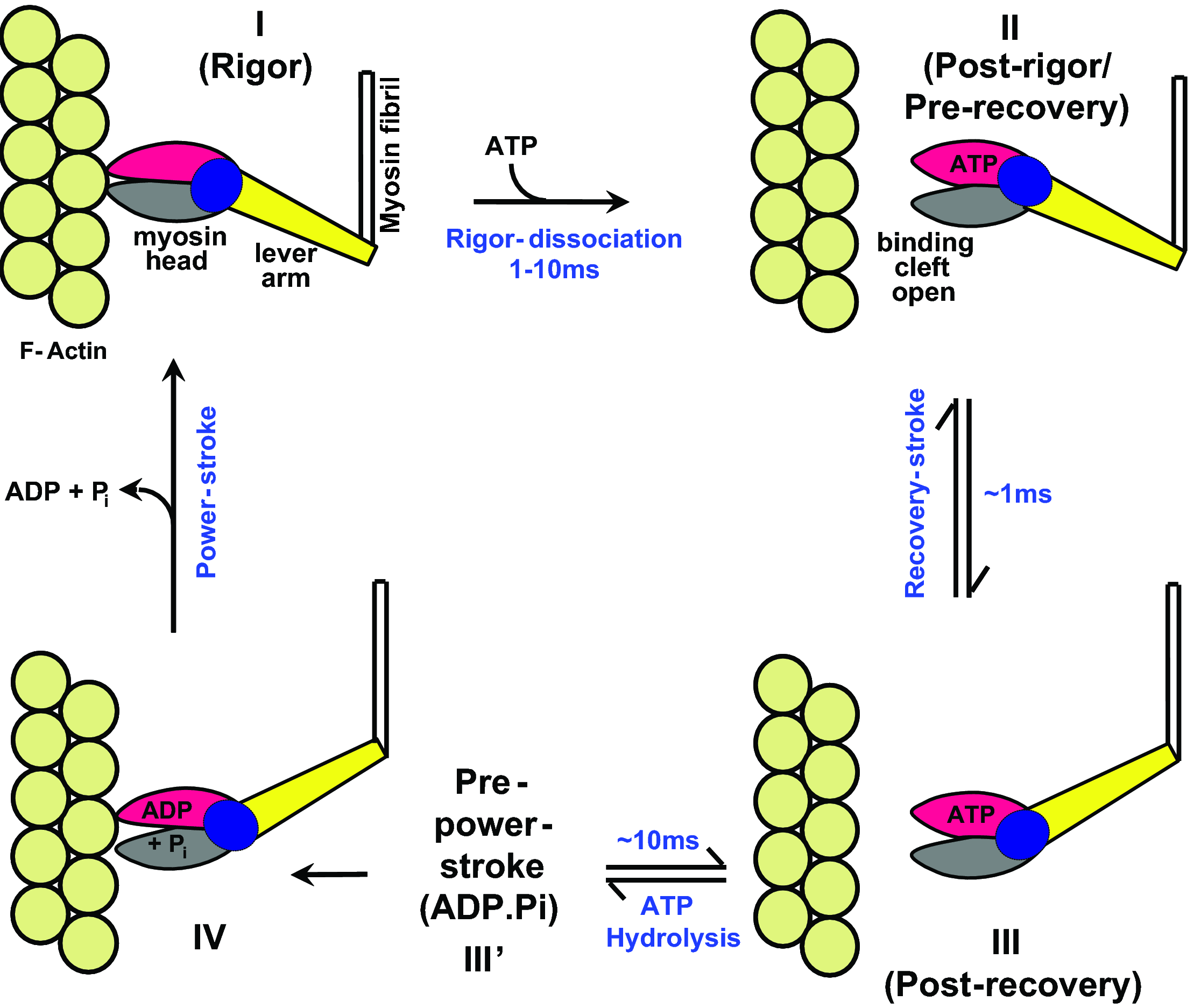
The
Lymn-Taylor cycle:
In the 'rigor dissociation'
step (I -> II), ATP binding to the myosin head in the 'rigor' state
(I) is coupled to dissociation from the F-actin fibril (olive spheres),
going to the post-rigor state (II).
Step II->III: Priming
(back swing) of the lever-arm (in yellow) is coupled to activation
of the ATPase.
Step III->III': ATP-hydrolysis
is coupled to an affinity-increase for actin.
Step III'->I: Rebinding to actin is coupled to the force-generating
swing of the lever arm.
|
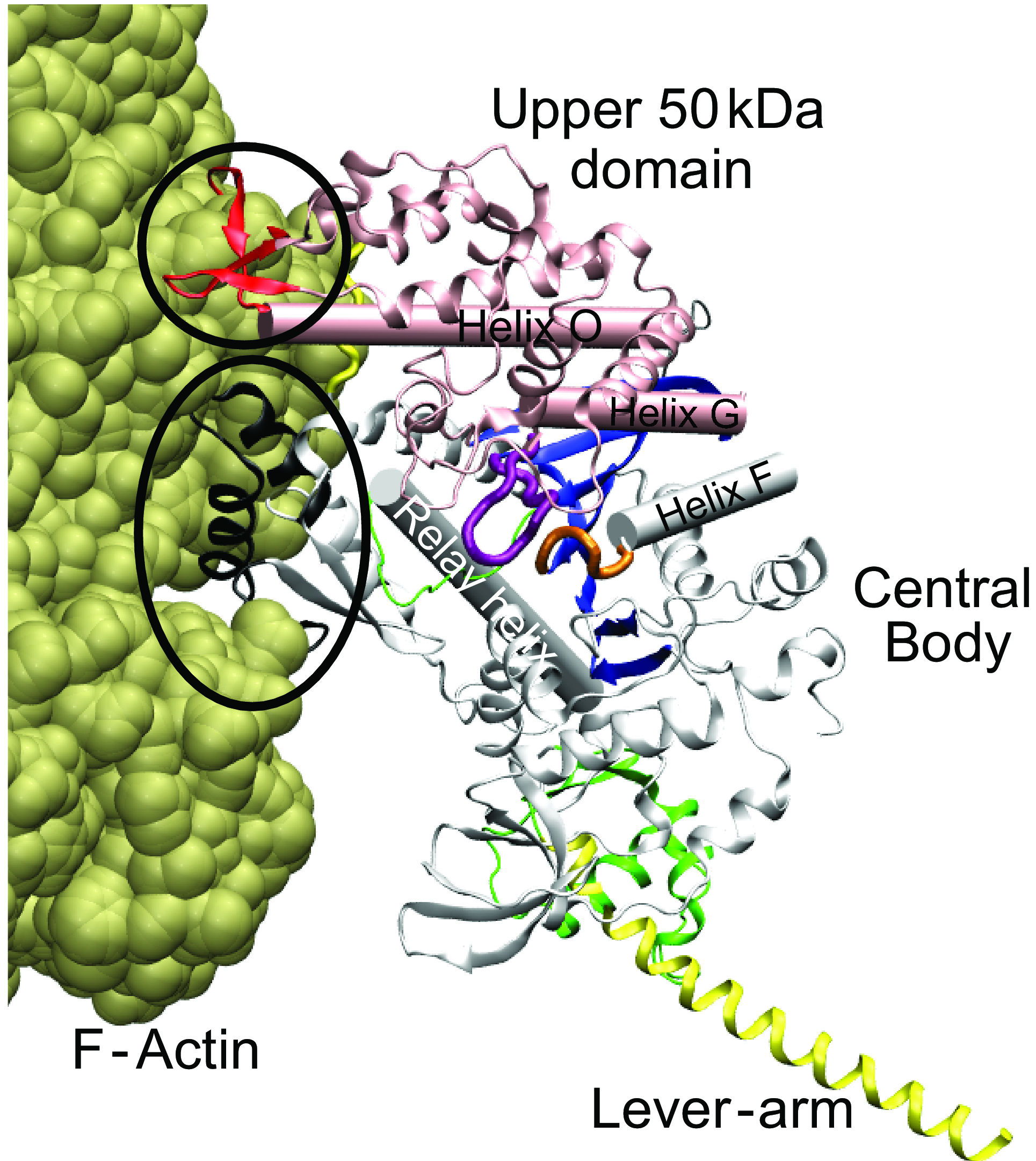
Rigor-like
conformation of the myosin head:
Shows the two regions (circled in black) that contact F-actin (in
olive). One region (in red) belongs to the upper 50kDa (u50) domain (in
pink), which is organized around helix-O. The other region (in black)
belongs to the lower 50 kDa domain (part of the central body, in grey).
These two domains are build around the central
ß-sheet (in blue) and are connected by a short strut (in yellow).
The ATP binding site is between the Switch-1 loop (in purple) and the
P-loop (in orange).
The converter domain is shown in thick green, the Switch-2 loop in thin
green. Same colours used throughout this page.
|
|
 |
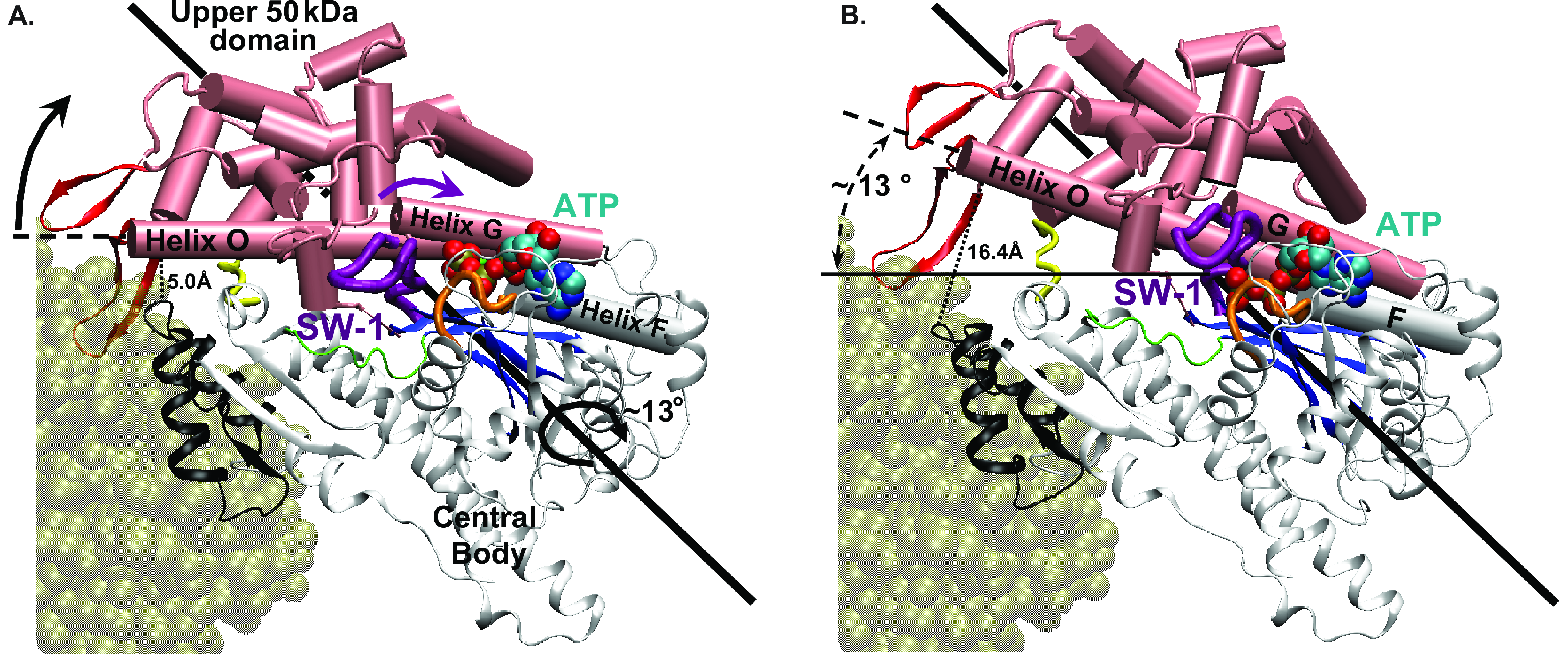
The transition pathway (Fig. 2 A & B of
paper):
Panel A shows the rigor state I, panel B
shows the post-rigor state II.
Thick arrows indicate motions during the transition
Movie 1 (showing the transition once):
The u50 domain (pink/red) and the attached Switch-1 loop (purple)
cohesively rotate by a 13° scissor motion relative to the central
body
(in grey/black), thereby twisting the central ß-sheet (in blue)
that
connects the u50 domain to the central body (the scissor axis is shown
as a thick black rod).
This opens the actin-binding cleft
(5->16.4Å), thus lowering the affinity for actin, and
sandwiches ATP
between the Switch-1 and the P- loops, thus stabilizing the post-rigor
state. F-actin (in olive, not present in the calcualtions) is
shown for orientation. The strut (in yellow) does not change its
length.
Movie 2 (showing the transition
back-and-forth, play with the "repeat" or "loop" option of your
video-player).
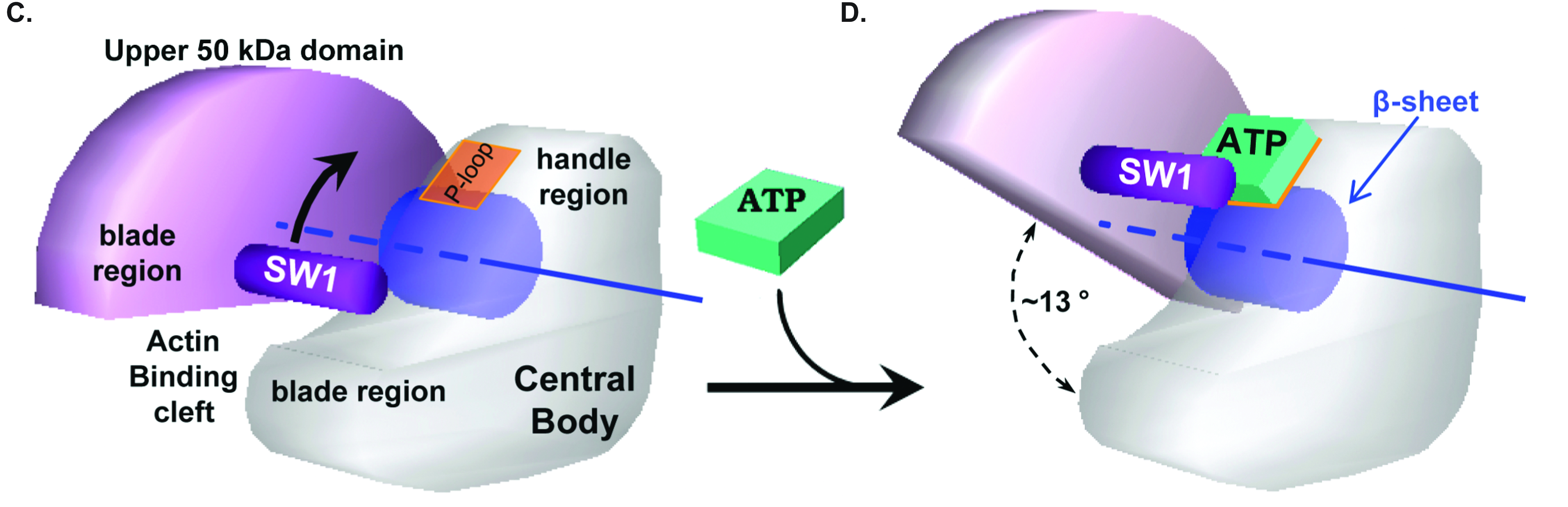
The mechanism (Fig. 2 C & D of the paper):
The present calculations reveal a simple coupling mechanism for the
ATP-induced rigor dissociation:
Movie 3, same as movie 2, but in
space-fill (back-and-forth, play with the "repeat" or "loop"
option of your video-player).
The u50 domain and the central body act as a pair of scissors which
pivot about an axis that passes through the middle of the central
ß-sheet (Panel C). The Switch-1 loop is attached to the
'scissor-blade' side of the u50 domain, while the P-loop is on the
'scissor-handle' side of the central body.
During the transition (Panel D), the scissors open (13°) by an
untwisting of the ß-sheet, bringing the Switch-1 loop close to
the P-loop (thereby increasing the affinity for ATP) and simultaneously
separating the two actin binding sites (thus lowering the affinity for
actin). This explains the reciprocal coupling between ATP and actin
binding.
The small change in distance between the P- and Switch-1 loops is
radially amplified into a large opening of the actin binding-cleft
because both loops are located near the pivot axis, while the two actin
binding sites are located at the tip of each blade (i.e., distant from
the pivot).
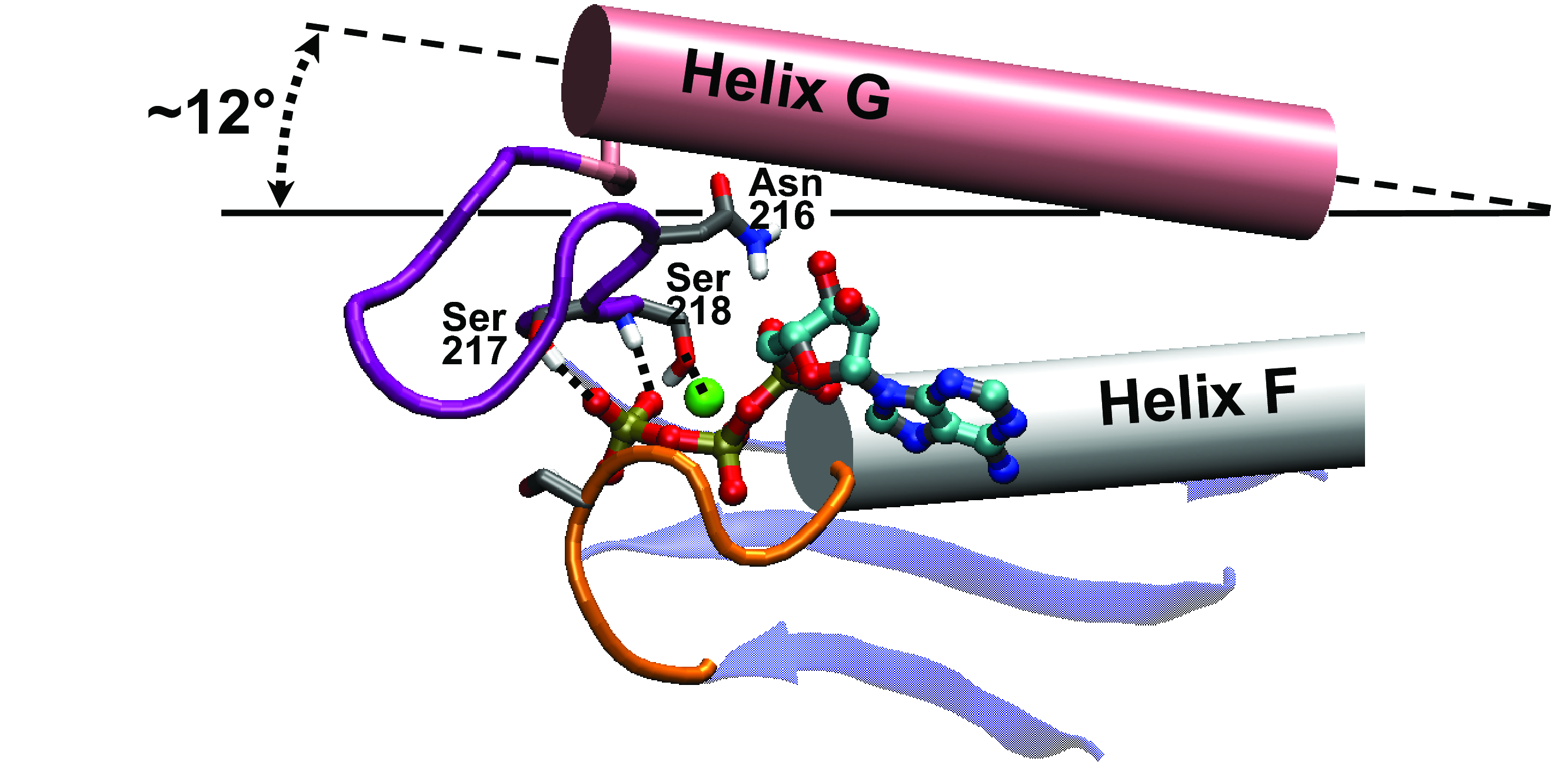
Interactions between Switch-1 and ATP along the
pathways:
The concerted rotation of helix G axis and Switch-1 loop (in
purple) accompany the
scissor motion of the u50 domain. Switch-1 and helix-G pivot by
12
degrees relative to helix F (belonging to the central body).
Switch-1 transiently interacts with the Mg/triphosphate (see
Supplemental Figure S2A) of ATP (phosphorus colored in gold).
In the final (post-rigor) state, the Switch-1
loop fully covers the triphosphate, and Ser217/218 forms key
interactions with the GAMMA-phosphate/Mg2+ (see Suppl. Fig. S2B).
Movie 4, starting with ATP already inside its pocket.
Movie 5, starting with ATP
placed outside its binding pocket.
For references, please see the PNAS
paper.