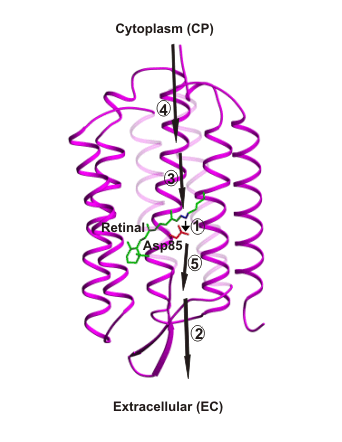
This figure shows the proton transfer steps in bacteriorhodopsin in the order they occur.
Step 1 is the "primary" transfer step, from the retinal Schiff-base NH to Asp85.
The three pathways with the lowest energy barrier are shown here:
Structure 12, p. 1281-1288 (2004). Full paper (PDF)
In the light-driven proton pump bacteriorhodopsin, after
photoisomerization
of the retinal chromophore, a proton is transferred from the retinal Schiff
base to the nearby Asp85.
Here, the mechanism of this transfer step is determined by computing
minimum-energy reaction paths in the protein using combined
quantum/classical
mechanics (QM/MM).
The results reveal that the Schiff base donor NH group first orients
in the direction opposite to that of the proton transfer. From this
stable
state, three mechanisms are found to have energy barriers in agreement
with experiment:
In one, the proton is transferred directly to Asp85. In the
second,
the proton is transferred along a proton wire via Thr89. In the
third,
the proton is transferred first to Asp212 and then to Asp85 along a
proton
wire via a water molecule.
Some of these pathways require considerable twisting of the Schiff
base accompanied by displacements of surrounding residues. Given the
close
proximity of the Schiff base and Asp85 (~4 Angstroem), the coexistence
of these very different transfer mechanisms is unexpected.
Alternative pathways
in the presence of an additional water (which is hydrogen bonded to
retinal on the cytoplasmic side) are also energetically allowed.
| The absorption of one photon by the all-trans
retinal chromophore
leads to rotation of the C13-C14 bond into the 13-cis conformation.
This
triggers a photocycle, the net effect of which is the transfer of one
proton
from the cytoplasmic to the extracellular side of the membrane.
This figure shows the proton transfer steps in bacteriorhodopsin in the order they occur. Step 1 is the "primary" transfer step, from the retinal Schiff-base NH to Asp85. |
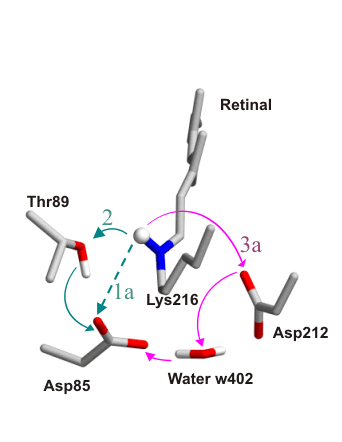
Several mechanisms are conceivable for primary proton
transfer, shown
here : The proton transfers could take place on either side of
retinal
(green and magenta arrows, respectively) and could be either direct
(broken
lines), or involve intermediate proton carriers and/or proceed via a
proton
wire (continuous lines). An exhaustive exploration of these different
mechanisms
was performed by computing all possible reaction pathways and their
transition
states with the Conjugate Peak Refinement
(CPR) method. The three pathways with the lowest energy barrier are shown here: |
|
|
Path 1a.
Download
the movie (1.2Mb)
Direct proton transfer from the retinal Schiff base to Asp85.
The shortening of the donor-acceptor distance involves significant
twisting of the retinal chain and simultaneous displacement of the
Asp85
side chain. Rate-limiting barrrier 12.4kcal/mol.
Path 2.
Download
the movie (0.5Mb)
The Schiff base proton is transferred to the hydroxyl group of Thr89,
whose own proton is transferred to Asp85 in a concerted manner.
The
initial hydrogen bonds are optimally arranged for this proton wire,
such
that little change in retinal twisting is needed, which distinguishes
this
mechanism from the others. Rate-limiting barrrier 13.6kcal/mol.
Path 3a.
Proton transfer to Asp212 followed by proton wire through water w402.
This pathway begins with a conformational change in retinal twisting: Download the movie (1.1Mb)
The subsequent proton transfer steps take place with little
structural
rearrangement. First, the Schiff base proton is transferred to Asp212.
The hydrogen-bonding pattern around Asp212 then reorganizes to form a
proton
wire from Asp212 to Asp85 via water w402. Rate-limiting barrrier
11.5kcal/mol.
Download the movie (2.4Mb)
Conclusions
Mechanisms 1a and 3a require extensive flexibility in
the retinal chain and neighbouring residues to bring the donor-acceptor
distance within proton transfer range. Because these proton
transfer mechanisms involve the complex interplay
of many different degrees of freedom in addition to proton
displacement,
such as retinal twisting and water rotation, the 'a priori' definition
of a suitable reaction coordinate is not practical and the use of an
automated
method like CPR for finding the minimum
energy
paths is indispensable. The usefulness of the QM/MM approach
is clear in this case, by allowing the donor and acceptor groups of the
quantum region to be carried flexibly by the protein scaffold.